
Imunoglobulinas (Ig) são Moléculas de glicoproteína que são produzidas pelos plasmócitos em resposta a um imunógeno e que funcionam como anticorpos. As imunoglobulinas derivam seu nome da descoberta de que elas migram com as proteínas globulares quando soro contendo anticorpos é colocado em um campo elétrico.


FUNÇÕES GERAIS DAS IMUNOGLOBULINAS
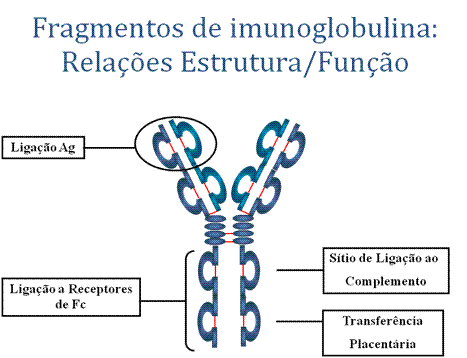
A. Ligação a antígeno
Imunoglobulinas se ligam especificamente a um ou a alguns antígenos proximamente relacionados. Cada imunoglobulina na verdade liga-se a um determinante antigênico específico. Ligação a antígeno pelos anticorpos é a função primária dos anticorpos e pode resultar em proteção do hospedeiro. A valência do anticorpo refere-se ao número de determinantes antigênicos que uma molécula individual de anticorpo pode se ligar. A valência de todos os anticorpos é pelo menos duas e em alguns casos mais.
B. Funções Efetoras
Freqüentemente a ligação de um anticorpo a um antígeno não tem efeito biológico direto. Ao invés disso, os efeitos biológicos significantes são uma conseqüência de “funções efetoras” secundárias de anticorpos. As imunoglobulinas mediam uma variedade dessas funções efetoras. Usualmente a habilidade de carrear uma função efetora particular requer que o anticorpo se ligue a seu antígeno. Nem todas as imunoglobulinas irão mediar todas as funções efetoras. Tais funções efetoras incluem:1. Fixação ao complemento – Isso resulta na lise de células e liberação de moléculas biologicamente ativas;
2. Ligação a vários tipos celulares – Células fagocitárias, linfócitos, plaquetas, células master, e basófilos têm receptores que se ligam a imunoglobulinas. Essa ligação pode ativar as células que passam a realizar algumas funções. Algumas imunoglobulinas também se ligam a receptores em trofoblastos placentários, o que resulta na transferência da imunoglobulina através da placenta. Como resultado, os anticorpos maternos transferidos provêem imunidade ao feto e ao recém-nascido.
ESTRUTURA BÁSICA DAS IMUNOGLOBULINAS
Embora diferentes imunoglobulinas possam diferir estruturalmente elas são todas construidas a partir das mesmas unidades básicas.
A. Cadeias leves e Pesadas Todas as imunoglobulinas têm uma estrutura de quatro cadeias como unidade básica. Elas são compostas de duas cadeias leves idênticas (23kD) e duas cadeias pesadas idênticas (50-70kD).
B. Pontes dissulfeto
C. Regiões Variáveis (V) e Constantes (C)1. Pontes dissulfeto intercadeia – As cadeias pesada e leve e as duas cadeias pesadas são mantidas juntas por pontes dissulfeto intercadeia e por interações não covalentes. O número de pontes dissulfeto varia entre as diferentes moléculas de imunoglobulinas.
2. Pontes dissulfeto intracadeia – Dentro de cada uma das cadeias polipeptídicas há também pontes dissulfeto intracadeia.
Depois que as sequências de aminoácidos de muitas cadeias pesadas e leves diferentes foram comparadas, ficou claro que ambas as cadeias pesadas e leves poderiam ser divididas em duas regiões baseando-se na variabilidade da seqüência de aminoácidos. Elas são:
D. Região da dobradiça1. Cadeia leve - VL (110 aminoácidos) e CL (110 aminoácidos)
2. Cadeia Pesada - VH (110 aminoácidos) e CH (330-440 aminoácidos)
Esta é a região com a qual os braços da molécula de anticorpo formam um Y. É chamada de região da dobradiça porque há uma flexibilidade na molécula nesse ponto.
E. Domínios
Imagens tridimensionais da molécula de imunoglobulina mostram que ela não é reta como mostrado na Figura. Ao contrário, ela é dobrada em regiões globulares, cada uma das quais contém uma ponte dissulfeto intracadeia. Essas regiões são chamadas domínios.
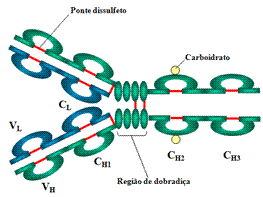
1. Domínios de Cadeia Leve - VL e CL
2. Domínios de Cadeia Pesada - VH, CH1 - CH3 (ou CH4)
F. Oligosacarídeos
Carboidratos são acoplados ao domínio CH2 na maioria das imunoglobulinas. Entretanto, em alguns casos carboidratos podem também serem acoplados em outros locais.
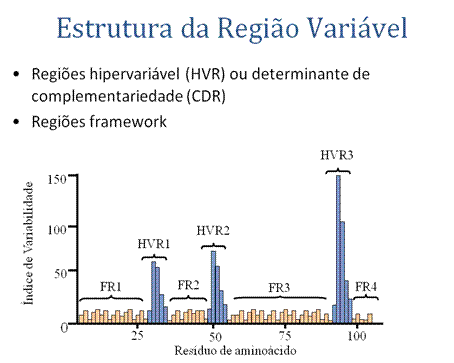
ESTRUTURA DA REGIÃO VARIÁVEL
A. Regiões hipervariáveis (HVR) ou regiões determinadoras de complementaridade (CDR)
B. Regiões frameworkComparações entre as seqüências de aminoácidos das regiões variáveis das imunoglobulinas mostram que a maioria das variações reside em três regiões chamadas de regiões hipervariáveis ou regiões determinadoras de complementaridade, como ilustrado na Figura . Anticorpos com especificidades diferentes (i.e. diferentes sítios de combinação) têm diferentes regiões determinadoras de complementariedade, enquanto que anticorpos de exatamente mesma especificidade têm regiões determinadoras de complementariedade idênticas (i.e. CDR é o sítio de combinação do anticorpo). Regiões determinadoras de complementariedade são encontradas em ambas as cadeias H e L.
As regiões entre as regiões determinadoras de complementariedade na região variável são chamadas regiões framework . Com base nas similaridades e diferenças nas regiões framework as regiões variáveis da cadeia pesada e leve da imunoglobulina podem ser divididas em grupos e subgrupos. Estes representam os produtos de diferentes genes de região variável.
FRAGMENTOS DE IMUNOGLOBULINA: RELAÇÕES ESTRUTURA/FUNÇÃO
Fragmentos de imunoglobulinas produzidos por digestão proteolítica têm-se mostrado úteis na elucidação das relações de estrutura e função em imunoglobulinas.
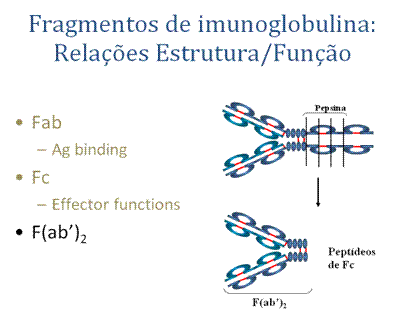
A. Fab
Digestão com papaína quebra a molécula de imunoglobulina na região da dobradiça antes da ponte dissulfeto intercadeia Figura . Isso resulta na formação de dois fragmentos idênticos que contém a cadeia leve e os domínios VH e CH1 da cadeia pesada.Ligação a antígeno – Esses fragmentos foram chamados de fragmentos Fab porque eles continham o sítio de ligação a antígenos do anticorpo. Cada fragmento Fab é monovalente enquanto que a molécula original era divalente. O sítio de combinação do anticorpo é criado tanto por VH e VL. Um anticorpo é capaz de se ligar a um determinante antigênico particular porque ele tem uma combinação particular de VH e VL. Combinações diferentes de VH e VL resultam em anticorpos capazes de se ligar a determinantes antigênicos diferentes.

B. Fc
Digestão com papaína também produz um fragmento que contém o restante das duas cadeias pesadas, cada uma contendo um domínio CH2 e CH3. Esse fragmento foi chamado Fc porque é facilmente cristalizado.
 Funções efetoras – As funções efetoras das imunoglobulinas são mediadas por esta parte da molécula. Diferentes funções são mediadas por diferentes domínios nesse fragmento. Normalmente a habilidade de um anticorpo exercer uma função efetora requer a ligação prévia a um antígeno; entretanto, há exceções a essa regra.
Funções efetoras – As funções efetoras das imunoglobulinas são mediadas por esta parte da molécula. Diferentes funções são mediadas por diferentes domínios nesse fragmento. Normalmente a habilidade de um anticorpo exercer uma função efetora requer a ligação prévia a um antígeno; entretanto, há exceções a essa regra.
C. F(ab')2
Tratamento de imunoglobulinas com pepsina resulta na clivagem da cadeia pesada depois das pontes dissulfeto H-H intercadeia, resultando em um fagmento que contém ambos os sítios de ligação a antígenos. Esse fragmento foi chamado F(ab')2 porque é divalente. A região Fc da molécula é digerida a pequenos peptídeos pela pepsina. O F(ab')2 liga-se a antígeno mas não media as funções efetoras dos anticorpos.A. Classes de imunoglobulinas
As imunoglobulinas podem ser divididas em cinco classes diferentes, com base nas diferenças em seqüências de aminoácidos na região constante das cadeias pesadas. Todas a imunoglobulinas de uma mesma classe tem regiões constantes de cadeia pesada muito similares. Essas diferenças podem ser detectadas por estudos de seqüências ou mais comumente por meios sorológicos (i.e. pelo uso de anticorpos dirigidos a essas diferenças).
1. IgG – Cadeias pesadas gama
2. IgM - Cadeias pesadas mu
3. IgA - Cadeias pesadas alfa
4. IgD - Cadeias pesadas delta
5. IgE - Cadeias pesadas épsilon
B. Subclasses de imunoglobulinas
As classes de imunoglobulinas podem ser divididas em subclasses baseadas em pequenas diferenças nas seqüências de aminoácidos na região constante das cadeias pesadas. Todas as imunoglobulinas de uma subclasse têm seqüências de aminoácidos de região constante de cadeia pesada muito similares. Novamente essas diferenças são mais comumente detectadas por meios sorológicos.
1. Subclasses de IgG
a) IgG1 – Cadeias pesadas gama 12. Subclasses de IgA
b) IgG2 - Cadeias pesadas gama 2
c) IgG3 - Cadeias pesadas gama 3
d) IgG4 - Cadeias pesadas gama 4
a) IgA1 - Cadeias pesadas alfa 1
b) IgA2 - Cadeias pesadas alfa 2
C. Tipos de imunoglobulinas
Imunoglogulinas podem ser também classificadas pelo tipo de cadeia leve que possuem. Tipos de cadeia leve são baseados na diferença de sequência de aminoácidos na região constante da cadeia leve. Essas diferenças são detectadas por meios sorológicos.
1. Cadeias leves kappa
2. Cadeias leves lambda
D. Subtipos de imunoglobulina
As cadeias leves podem ser também divididas em subtipos baseados nas diferenças de sequência de aminoácidos da região constante de cadeia leve.
1. Subtipos de lambda
a) Lambda 1
b) Lambda 2
c) Lambda 3
d) Lambda 4
E. Nomenclatura
Imunoglobulinas são nomeadas com base na classe, ou na cubclasse de cadeia pesada e tipo ou subtipo de cadeia leve. A menos que seja precisamente declarado você deve assumir que todas as subclasses, tipos e subtipos estão presentes. IgG significa que todas as subclasses e tipos estão presentes.
F. Heterogeneidade
Imunoglobulinas consideradas como uma população de moléculas são normalmente muito heterogêneas porque elas são compostas de diferentes classes e subclasses cada uma com diferentes tipos e subtipos de cadeias leves. Além disso, diferentes moléculas de imunoglobulinas podem ter diferentes propriedades de ligação a antígenos devido às diferentes regiões VH e VL.
ESTRUTURA E ALGUMAS PROPRIEDADES DE CLASSES E SUBCLASSES DE IG
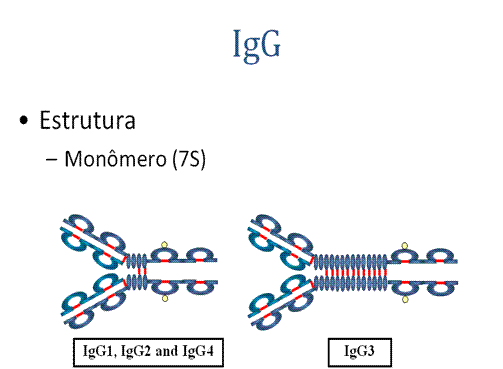
A. IgG
1. EstruturaAs estruturas de subclasses de IgG estão apresentadas na Figura . Todas IgG's são monômeros (imunoglobulina 7S). As subclasses diferem no número de pontes dissulfeto e comprimento da região da dobradiça.
2. Propriedades
A mais versátil imunoglobulina porque é capaz de realizar todas as funções das moléculas de imunoglobulinas.a) IgG é a principal Ig no soro - 75% das Ig do soro são IgG;
b) IgG é a principal Ig em espaços extra vasculares;
c) Transferência placentária - IgG é a única classe de Ig que atravessa a placenta. A transferência é mediada pelo receptor da região Fc do IgG nas células placentárias. Nem todas as subclasses atravessam com a mesma eficiência; IgG2 não atravessa bem;
d) Fixação do complemento – Nem todas as subclasses fixam com a mesma eficiência; IgG4 não fixa complemento;
e) Ligação a células – Macrófagos, monócitos, PMN's e alguns linfócitos têm receptores para a região Fc da IgG. Nem todas as subclasses se ligam com a mesma eficiência; IgG2 e IgG4 não se ligam a receptores de Fc. Uma consequência da ligação a receptores de Fc em PMN's, monócitos e macrófagos é que a célula pode então internalizar o antígeno melhor. O anticorpo preparou o antígeno para ser comido pelas células fagocitárias. O termo opsonina é usado para descrever substâncias que aumentam a fagocitose. IgG é uma boa opsonina. Ligação de IgG a receptores de Fc em outros tipos de células resulta na ativação de outras funções;
B. IgM
1. EstruturaA estrutura da IgM está apresentada na Figura. IgM normalmente existe como um pentâmero (imunoglobulina 19S) mas ela pode também existir como um monômero. Na forma pentamérica todas as cadeias pesadas são idênticas e todas as cadeias leves são idênticas. Assim, a valência é teoricamente 10. IgM tem um domínio extra na cadeia mu (CH4) e ela tem outra proteína covalentemente ligada via uma ponde S-S chamada cadeia J. Esta cadeia funciona em polimerização da molécula a um pentâmero.
2. Propiedadesa) IgM é a terceira Ig mais comum no soro.
b) IgM é a primeira Ig a ser feita pelo feto e a primeira Ig a ser feita por uma célula B virgem quando é estimulada pelo antígeno.
c) Como consequência da sua estrutura pentamérica, IgM é uma boa Ig fixadora do complemento. Assim, anticorpos IgM são muito eficientes em levar à lise de microrganismos.
d) Como consequência da sua estrutura, IgM também é uma boa Ig aglutinadora. Assim, anticorpos IgM são muito boas em agregar microrganismos para eliminação eventual para fora do corpo.
e) IgM liga-se a algumas células via receptores de Fc.
f) Ig de superfície de célula B
IgM de superfície existe como um monômero e não tem cadeia J mas tem 20 aminoácidos extras na região C-terminal para se ancorar na membrana (Figura 9). IgM de superfície celular funcionam como um receptor para antígeno ou células B. IgM de superfície é associada não covalentemente com duas proteínas adicionais na membrana da célula B chamadas Ig-alfa e Ig-beta como indicado na Figura 10. Essas proteínas adicionais agem como moléculas de transdução de sinal uma vez que a cauda citoplasmática da molécula de Ig por si mesma é muito curta para transduzir um sinal. O contato entre a superfície da imunoglobulina e um antígeno é necessário antes de um sinal ser transduzido pelas cadeias Ig-alfa e Ig-beta. No caso dos antígenos T-independentes, contato entre o antígeno e a superfície da imunoglobulina é suficiente para ativar as células B a se diferenciarem em plasmócitos secretores de anticorpos. Entretanto, para antígenos T-dependentes, um segundo sinal fornecido pelas células T auxiliares é necessário para ativar as células B.
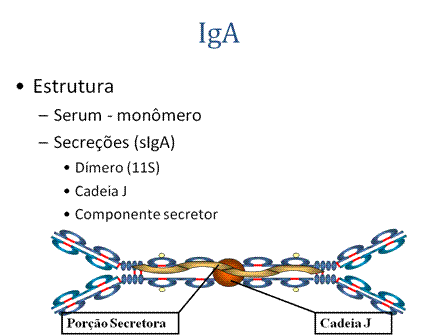
C. IgA
Quando IgA é encontrada em secreções também tem outra proteína associada a ela chamada de peça secretora T; sIgA é às vezes referida como imunoglobulina 11S. Ao contrário do resto da IgA que é feito no plasmócito, a peça secretora é feita nas células epiteliais e é adicionada à IgA à medida que esta passa através das secreções (Figura 12). A peça secretora ajuda a IgA a ser transportada através da mucosa e também a protege da degradação nas secreções.1. EstruturaA IgA do soro é um monômero mas a IgA encontrada em secreções é um dímero como apresentado na Figura. Quando IgA sai do dímero, uma cadeia J se associa a ela.
2. Propriedades
a) IgA é a 2a Ig mais comum no soro.b) IgA é a principal classe de Ig em secreções – lágrimas, saliva, colostro, muco. Uma vez que é encontrada em secreções IgA secretora é importante na imunidade local (de mucosa).c) Normalmente IgA não fixa complemento, a menos que esteja agregada.d) IgA pode se ligar a algumas células - PMN's e alguns linfócitos.
D. IgD
1. EstruturaA estrutura da IgD está apresentada na Figura . IgD existe somente como um monômero.
2. Propriedades
a) IgD é encontrada em baixos níveis no soro; seu papel no soro é duvidoso.
b) IgD primariamente encontrada em superfícies de célula B onde funciona como um receptor para antígeno. IgD na superfície de células B tem aminoácidos extras na região C-terminal para ancoramento à membrana. Ela também se associa com as cadeias beta de Ig-alfa e Ig-beta.
c) IgD liga complemento.E. IgE
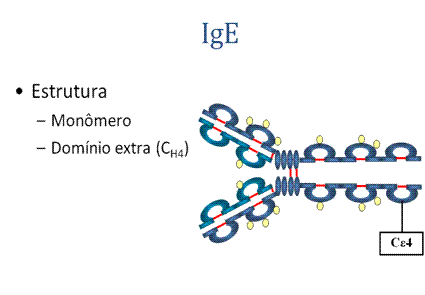
1. Estrutura
A estrutura do IgE está apresentada na Figura . IgE existe como um monômero e tem um domínio extra na região constante.
2. Propriedades
a) IgE é a Ig sérica menos comum uma vez que se liga fortemente com receptores de Fc em basófilos e mastócitos mesmo antes da interação com o antígeno.
b) Envolvida em reações alérgicas – Como consequência da sua ligação a basófilos e mastócitos, IgE é envolvida em reações alérgicas. Ligação do alergeno à IGe nas células resulta na liberação de vários mediadores farmacológicos que resulta em sintomas alérgicos.
c) IgE também participa em doenças parasitárias por helmintos. Uma vez que os níveis sorológicos de IgE aumentam em doenças parasitárias, a quantificação dos níveis de IgE auxilia no diagnóstico de infecções parasitárias. Eosinófilos têm receptores de Fc para IgE e a ligação de eosinófilos a helmintos cobertos por IgE resulta na morte do parasita.
d) IgE não fixa complemento.IMPLICAÇÕES CLÍNICAS DAS CLASSES DE IMUNOGLOBULINAS HUMANASIgG1. Aumenta em:
2. Diminui em:a) Infecções granulomatosas crônicas
b) Infecções de todos os tipos
c) Hiperimunização
d) Doenças hepáticas
e) Desnutrição (severa)
f) Disproteinemia
g) Doenças associadas com hipersensibilidade granulomas, desordens dermatológicas, e mieloma de IgG.
h) Artrite reumatóide
a) Agamaglobulinemia
b) Aplasia linfóide
c) Deficiência seletiva IgG, IgA
d) Mieloma de IgA
e) Proteinemia de Bence Jones
f) Leucemia linfoblástica crônicaIgM1. Aumenta (em adultos) em:
2. Diminui em:a) Macroglobulinemia de Waldenström
b) Tripanosomíase
c) Actinomicose
d) Doença de Carrión (bartonelose)
e) Malária
f) Mononucleose infecciosa
g) Lúpus eritematoso
h) Artrite reumatóide
I) Disgamaglobulinemia (certos casos)
Nota: No recém nascido, um nível de IgM superior a 20 ng./dl é uma indicação de estimulação do sistema imune in utero e estimulação pelo vírus da rubéola, citomegalovírus, sífilis, ou toxoplasmose.
a) Agamaglobulinemia
b) Desordens linfoproliferativas (certos casos)
c) Aplasia linfóide
d) Mieloma de IgG e IgA
e) Disgamaglobulinemia
f) Leucemia linfoblástica crônicaIgA1. Aumenta em:
a) Síndrome de Wiskott-Aldrich
b) Cirrose hepática (na maioria dos casos)
c) Certos estágios de desordens autoimunes do colágeno e outras, tais como artrite reumatóide e lúpus eritematoso
d) Infecções crônicas não baseadas em deficiências imunológicas
e) Mieloma de IgA2. Diminui em:a) Ataxia telangiectasia hereditária
b) Estados de deficiência imunológica (ex. disgamaglobulinemia, agamaglobulinemia congênita e adquirida, e hipogamaglobulinemia)
c) Síndromes de mal absorção
d) Aplasia linfóide
e) Mieloma de IgG
f) Leucemia linfoblástica aguda
g) Leucemia linfoblástica crônicaIgD1. Aumenta em:
a) Infecções crônicas
b) Mielomas de IgDIgE1. Aumenta em:
a) Doenças de pele atópicas tais como eczema
b) Febre do feno
c) Asthma
d) Choque anafilático
e) Mieloma de IgE
Fonte: Artigo Científico Dr. Gene Mayer / microbiologybook.org2. Diminui em:a) Agamaglobulinemia congênita
b) Hipogamaglobulinemia por defeito no metabolismo ou na síntese de imunoglobulinas.
Nenhum comentário:
Postar um comentário